However, the name change is not generally accepted by most researchers, who still publish Vibrio fischeri (see Google Scholar for 2018–2019).
Genome
The genome for A. fischeri was completely sequenced in 2004 and consists of two chromosomes, one smaller and one larger. Chromosome 1 has 2.9 million base pairs (Mbp) and chromosome 2 has 1.3 Mbp, bringing the total genome to 4.2 Mbp.
A. fischeri has the lowest G+C content of 27 Vibrio species, but is still most closely related to the higher-pathogenicity species such as V. cholerae. The genome for A. fischeri also carries mobile genetic elements.
A. fischeri are globally distributed in temperate and subtropicalmarine environments. They can be found free-floating in oceans, as well as associated with marine animals, sediment, and decaying matter. A. fischeri have been most studied as symbionts of marine animals, including squids in the genus Euprymna and Sepiola, where A. fischeri can be found in the squids’ light organs.
This relationship has been best characterized in the Hawaiian Bobtail Squid (Euprymna scolopes), where A. fischeri is the only species of bacteria inhabiting the squid’s light organ.
A. fischeri colonization of the light organ of the Hawaiian bobtail squid is currently studied as a simple model for mutualistic symbiosis, as it contains only two species and A. fischeri can be cultured in a lab and genetically modified. This mutualistic symbiosis functions primarily due to A. fischeri bioluminescence. A. fischeri colonizes the light organ of the Hawaiian bobtail squid and luminesces at night, providing the squid with counter-illumination camouflage, which prevents the squid from casting a shadow on the ocean floor.
Euprymna scolopes lives in a symbiotic relationship with the bioluminescentbacteriaAliivibrio fischeri, which inhabits a special light organ in the squid’s mantle. The bacteria are fed a sugar and amino acid solution by the squid and in return hide the squid’s silhouette when viewed from below by matching the amount of light hitting the top of the mantle (counter-illumination). E. scolopes serves as a model organism for animal-bacterial symbiosis and its relationship with A. fischeri has been carefully studied.
Young, R.E. & C.F. Roper 1976. Bioluminescent countershading in midwater animals: evidence from living squid. Science191(4231): 1046–1048. doi:10.1126/science.1251214
The bioluminescent bacterium, A. fischeri, is horizontally transmitted throughout the E. scolopes population. Hatchlings lack these necessary bacteria and must carefully select for them in a marine world saturated with other microorganisms.
To effectively capture these cells, E. scolopes secretes mucus in response to peptidoglycan (a major cell wall component of bacteria).The mucus inundates the ciliated fields in the immediate area around the six pores of the light organ and captures a large variety of bacteria. However, by some unknown mechanism, A. fischeri is able to outcompete other bacteria in the mucus.
The evolutionary ecology of a sepiolid squid-Aliivibrio association: from cell to environment. Vie et Milieu58(2): 175-184. ISSN0240-8759
As A. fischeri cells aggregate in the mucus, they must use their flagella to migrate through the pores and down into the ciliated ducts of the light organ and endure another barrage of host factors meant to ensure only A. fischeri colonization. Besides the relentless host-derived current that forces motility-challenged bacteria out of the pores, a number of reactive oxygen species makes the environment unbearable. Squid halide peroxidase is the main enzyme responsible for crafting this microbiocidal environment, using hydrogen peroxide as a substrate, but A. fischeri has evolved a brilliant counterattack. A. fischeri possesses a periplasmic catalase that captures hydrogen peroxide before it can be used by the squid halide peroxidase, thus inhibiting the enzyme indirectly. Once through these ciliated ducts, A. fischeri cells swim on towards the antechamber, a large epithelial-lined space, and colonize the narrow epithelial crypts.
The evolutionary ecology of a sepiolid squid-Aliivibrio association: from cell to environment. Vie et Milieu58(2): 175-184. ISSN0240-8759
The bacteria thrive on the host-derived amino acids and sugars in the antechamber and quickly fill the crypt spaces within 10 to 12 hours after hatching.
An exclusive contract: Specificity in the Aliivibrio fischeriEuprymna scolopes partnership. Journal of Bacteriology182(7): 1779-1787. ISSN0021-9193
Ongoing relationship
Every second, a juvenile squid ventilates about 2.6 ml (0.092 imp fl oz; 0.088 US fl oz) of ambient seawater through its mantle cavity. Only a single A. fischeri cell, one/1-millionth of the total volume, is present with each ventilation.
The evolutionary ecology of a sepiolid squid-Aliivibrio association: from cell to environment. Vie et Milieu58(2): 175-184. ISSN0240-8759
The increased amino acids and sugars feed the metabolically demanding bioluminescence of the A. fischeri, and in 12 hours, the bioluminescence peaks and the juvenile squid is able to counterilluminate less than a day after hatching. Bioluminescence demands a substantial amount of energy from a bacterial cell. It is estimated to demand 20% of a cell’s metabolic potential.
An exclusive contract: Specificity in the Aliivibrio fischeriEuprymna scolopes partnership. Journal of Bacteriology182(7): 1779-1787. ISSN0021-9193
Nonluminescent strains of A. fischeri would have a definite competitive advantage over the luminescent wild-type, however nonluminescent mutants are never found in the light organ of the E. scolopes. In fact, experimental procedures have shown that removing the genes responsible for light production in A. fischeri drastically reduces colonization efficiency. Luminescent cells, with functioning luciferase, may have a higher affinity for oxygen than for peroxidases, thereby negating the toxic effects of the peroxidases. For this reason, bioluminescence is thought to have evolved as an ancient oxygen detoxification mechanism in bacteria.
An exclusive contract: Specificity in the Aliivibrio fischeriEuprymna scolopes partnership. Journal of Bacteriology182(7): 1779-1787. ISSN0021-9193
A. fischeri colonization occurs in juvenile squids and induces morphological changes in the squids light organ. Interestingly, certain morphological changes made by A. fischeri do not occur when the microbe cannot luminesce, indicating that bioluminescence (described below) is truly essential for symbiosis. In the process of colonization, ciliated cells within the animals’ photophores (light-producing organs) selectively draw in the symbiotic bacteria. These cells promote the growth of the symbionts and actively reject any competitors. The bacteria cause these cells to die off once the light organ is sufficiently colonized.
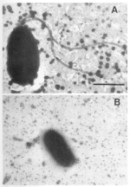
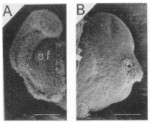
When a juvenile squid hatches from the egg, it does not contain any symbionts (it is aposymbiotic). It needs to acquire the symbionts from the sea water before it can use its light organ. The light organ of such a hatchling has modifications that apparently aid the hatchlingin obtaining the symbionts from the multitude of bacteria present in sea water. The most obvious modification are ciliated “arms” that circulate sea water over the pores of the empty light organ crypts (right picture, panel A shows one lobe of the bilobed juvenile light organ). Powered by their flagella (left picture, panel A), motile V. fischeri enter the pores of the light organ, move into the empty crypts and begin to grow rapidly. The presence of the symbionts influences the development of the host. The ciliated arms regress by apoptosis (right picture, panel B) and the bacteria are packed tightly in the crypts (picture below on the right, the arrow points at the symbionts inside the crypt that is lined by epithelial cells). Several hours after the bacteria have entered the light organ, the symbionts change; they loose their flagella, decrease in size and begin to emit light (left picture, panel B). Within a few weeks after the bacteria colonize the squid, the fully developed light organ is present. The light organ possess a silver-colored reflector tissue, a shutter mechanism (the black ink sack), a transparent lens that covers the light organ and a yellow filter that changes the color of the emitted light (shown below left). This allows the squid to control the amount of light that it emits.
J.Graf The Light-Organ Symbiosis of Vibrio fischeri and the Hawaiian squid, Euprymna scolopes, 2005, Wayback Machine https://web.archive.org/web/20070426122929 http://web.uconn.edu/mcbstaff/graf/VfEs/VfEssym.htm
In September 2002, the genome sequence of V. fischeri became available to the public. The strain that was sequenced, ES114, was isolated from the light organ of E. scolopes in 1988. For more information visit the home page of the Vibrio fischeri genome project.
The mutants that have been tested so far can be grouped into five different classes: (1) initiation mutants, these are mutants that cannot colonize the light organ to a detectable level; (2) accommodation mutants, these are mutants that can grow inside the light organ, but do not reach the same level of colonization as the wild type strain; (3) persistence mutants colonize as well as the wild type strain until after a certain time point they begin to decrease in number; (4) competition mutants, these mutants can colonize and persist just as well as the wild type strain when the mutant strain enters the light organ alone, but if the wild type strain is also present, the mutant is outcompeted by the wild type strain and does not reach its normal level of colonization; and (5) none, no defect in symbiotic association was detected using the standard conditions.
The following mutants have been tested for symbiotic competence (modified from Ruby, 1996).
Symbionts and host influence each others development.
The light organs of certain squid contain reflective plates that intensify and direct the light produced, due to proteins known as reflectins. They regulate the light for counter-illuminationcamouflage, requiring the intensity to match that of the sea surface above. Sepiolid squid expel 90% of the symbiotic bacteria in their light organ each morning in a process known as “venting”. Venting is thought to provide the source from which newly hatched squid are colonized by A. fischeri.
The light organ has an electrical response when stimulated by light, which suggests the organ functions as a photoreceptor that enables the host squid to respond to A. fischeri’s luminescence.
Tong, D., N.S. Rozas, T.H. Oakley, J. Mitchell, N.J. Colley & M.J. McFall-Ngai 2009. Evidence for light perception in a bioluminescent organ. PNAS106(24): 9836–9841. doi:10.1073/pnas.0904571106
Extraocular vesicles collaborate with the eyes to monitor the down-welling light and light created from counterillumination, so as the squid moves to various depths, it can maintain the proper level of output light. Acting on this information, the squid can then adjust the intensity of the bioluminescence by modifying the ink sac, which functions as a diaphragm around the light organ. Furthermore, the light organ contains a network of unique reflector and lens tissues that help reflect and focus the light ventrally through the mantle.
The light organ of embryonic and juvenile squids has a striking anatomical similarity to an eye and expresses several genes similar to those involved in eye development in mammalian embryos (e.g. eya, dac) which indicate that squid eyes and squid light organs may be formed using the same developmental “toolkit”.
Madigan M, Martinko J, eds. (2005). Brock Biology of Microorganisms (11th ed.). Prentice Hall. ISBN978-0-13-144329-7.
The population of A. fischeri needs to reach an optimal level to activate the lux operon and stimulate light production. The circadian rhythm controls light expression, where luminescence is much brighter during the day and dimmer at night, as required for camouflage.
The bacterial luciferin–luciferase system is encoded by a set of genes labelled the lux operon. In A. fischeri, five such genes (luxCDABEG) have been identified as active in the emission of visible light, and two genes (luxR and luxI) are involved in regulating the operon. Several external and intrinsic factors appear to either induce or inhibit the transcription of this gene set and produce or suppress light emission.
A. fischeri is one of many species of bacteria that commonly form symbiotic relationships with marine organisms. Marine organisms contain bacteria that use bioluminescence so they can find mates, ward off predators, attract prey, or communicate with other organisms. In return, the organism the bacteria are living within provides the bacteria with a nutrient-rich environment.
The lux operon is a 9-kilobase fragment of the A. fischeri genome that controls bioluminescence through the catalytic activity of the enzyme luciferase. This operon has a known gene sequence of luxCDAB(F)E, where luxA and luxB code for the protein subunits of the luciferase enzyme, and the luxCDE codes for a fatty acid reductase complex that makes the fatty acids necessary for the luciferase mechanism. luxC codes for the enzyme acyl-reductase, luxD codes for acyl-transferase, and luxE makes the proteins needed for the enzyme acyl-protein synthetase. Luciferase produces blue/green light through the oxidation of reduced flavin mononucleotide and a long-chain aldehyde by diatomic oxygen.
(Meighen, 1991).
Silverman et al., 1984
The reaction is summarized as:
FMNH2 + O2 + R-CHO → FMN + R-COOH + H2O + light.
The reduced flavin mononucleotide (FMNH) is provided by the fre gene, also referred to as luxG. In A. fischeri, it is directly next to luxE (giving luxCDABE-fre) from 1042306 to 1048745
To generate the aldehyde needed in the reaction above, three additional enzymes are needed. The fatty acids needed for the reaction are pulled from the fatty acid biosynthesis pathway by acyl-transferase. Acyl-transferase reacts with acyl-ACP to release R-COOH, a free fatty acid. R-COOH is reduced by a two-enzyme system to an aldehyde.
Quorum sensing in Aliivibrio fischeri[16]Green pentagons denote AHL autoinducer that LuxI produces (3OC6-homoserine lactone). Transcriptional regulator, LuxR, modulates expression of AHL synthase, LuxI, and the lux operon, leading to luciferase-mediated light emission
One primary system that controls bioluminescence through regulation of the luxoperon is quorum sensing, a conserved system across many microbial species that regulates gene expression in response to bacterial concentration. Quorum sensing functions through the production of an autoinducer, usually a small organic molecule, by individual cells. As cell populations increase, levels of autoinducers increase, and specific proteins that regulate transcription of genes bind to these autoinducers and alter gene expression. This system allows microbial cells to “communicate” amongst each other and coordinate behaviors like luminescence, which require large amounts of cells to produce an effect.
Waters, Christopher M.; Bassler, Bonnie L. (2005). “Quorum sensing: Cell-to-cell communication in bacteria”. Annual Review of Cell and Developmental Biology. 21: 319–346. doi:10.1146/annurev.cellbio.21.012704.131001. PMID16212498
In A. fischeri, there are two primary quorum sensing systems, each of which respond to slightly different environments. The first system is commonly referred to as the lux system, as it is encoded within the lux operon, and uses the autoinducer 3OC6-HSL. The protein LuxI synthesizes this signal, which is subsequently released from the cell. This signal, 3OC6-HSL, then binds to the protein LuxR, which regulates the expression of many different genes, but is most known for upregulation of genes involved in luminescence. The second system, commonly referred to as the ain system, uses the autoinducer C8-HSL, which is produced by the protein AinS. Similar to the lux system, the autoinducer C8-HSL increases activation of LuxR. In addition, C8-HSL binds to another transcriptional regulator, LitR, giving the ain and lux systems of quorum sensing slightly different genetic targets within the cell.
The different genetic targets of the ain and lux systems are essential, because these two systems respond to different cellular environments. The ain system regulates transcription in response to intermediate cell density cell environments, producing lower levels of luminescence and even regulating metabolic processes like the acetate switch.
On the other hand, the lux quorum sensing system occurs in response to high cell density, producing high levels of luminescence and regulating the transcription of other genes, including QsrP, RibB, and AcfA.
Activation of the lux operon by LuxR and LuxI in Aliivibrio fischeri[22][23](A) At low cell density, the autoinducers (3OC6-HSL – red dots), produced by LuxI, diffuse through the cell membrane into the growth medium (B) As the cell growth continues, the autoinducers in the medium start to accumulate in a confined environment. A very low intensity of light can be detected. (C) When enough autoinducers have accumulated in the medium, they can re-enter the cell where they directly bind the LuxR protein to activate luxICDABEG expression. (D) High levels of autoinducers activate the luminescent system of A. fischeri. A high intensity of light can be detected.
Natural transformation
Natural bacterial transformation is an adaptation for transferring DNA from one individual cell to another. Natural transformation, including the uptake and incorporation of exogenous DNA into the recipient genome, has been demonstrated in A. fischeri.
This process requires induction by chitohexaose and is likely regulated by genes tfoX and tfoY. Natural transformation of A. fischeri facilitates rapid transfer of mutant genes across strains and provides a useful tool for experimental genetic manipulation in this species.
On June 3, 2021, SpaceX CRS-22 launched E. scolopes, along with tardigrades, to the International Space Station. The squid were launched as hatchlings and will be studied to see if they can incorporate their symbiotic bacteria into their light organ while in space.
Waters, Christopher M.; Bassler, Bonnie L. (2005). “Quorum sensing: Cell-to-cell communication in bacteria”. Annual Review of Cell and Developmental Biology. 21: 319–346. doi:10.1146/annurev.cellbio.21.012704.131001. PMID16212498.
Visick, K.V. and M.J. McFall-Ngai. 2000. An exclusive contract: specificity in the Vibrio fischeri–Euprymna scolopes partnership. J. Bacteriol. 182:1779-1787.
McFall-Ngai, M.J. 1999. Consequences of evolution with bacterial symbionts: insights from the squid-Vibrio association. Annu. Rev. Ecol. Syst. 30:235-256.
Ruby, E. G. 1999. Ecology of a benign “infection”: colonization of the squid luminous organ by Vibrio fischeri. p. 217-231. In: E. Rosenberg (ed.), Microbial ecology and infectious disease. ASM Press, Washington, D.C.
Ruby, E. G. and M. J. McFall-Ngai. 1999. Oxygen-utilizing reactions and symbiotic colonization of the squid light organ by Vibrio fischeri. Trends Microbiol. 7:414-20.
McFall-Ngai, M.J., and E.G. Ruby. 1998. Bobtail squid and their luminous bacteria: when first they meet. BioScience 48:257-265.
Ruby, E.G., and K.-H. Lee. 1998. The Vibrio fischeri-Euprymna scolopes light organ association: current ecological paradigms. Appl. Environ. Microbiol. 64:805-812.
Ruby, E.G. 1996. Development of a cooperative, bacterial-animal symbiosis: the Vibrio fischeri-Euprymna scolopes light organ symbiosis. Annu. Rev. Microbiol. 50:591-624.
Koropatnick, T. A., J. T. Engle, M.A. Apicella, E. V. Stabb, W. E. Goldman, M.J. McFall-Ngai. 2004. Microbial factor-mediated development in a host-bacterial mutualism. Science. 306:1186-1188.
Kimbell, J., and M.J. McFall-Ngai. 2004. Symbiont-induced changes in host actin during the onset of a beneficial animal-bacterial association. Appl. Environ. Microbiol. 70:1434-1441.
DeLooney-Marino, C.R., A. J. Wolfe, K. L. Visick. 2003. Chemoattraction in Vibrio fischeri to serine, nucleosides, and N-acetylneuraminic acid, a component of squid light-organ mucous. Appl. Environ. Microbiol. 69:7527-7530.
Nishiguchi, M. K. and V. S. Nair. Evolution of symbiosis in the Vibrionaceae: a combined approach using molecules and physiology. Int. J. Syst. Evol. Microbiol. 54:2019-2026.
Fidopiastis PM, Miyamoto CM, Jobling MG, Meighen EA, Ruby EG. 2002. LitR, a new transcriptional activator in Vibrio fischeri, regulates luminescence and symbiotic light organ colonization. Mol. Microbiol. 45:131-43.
Millikan, D. S. and E. G. Ruby. 2002. Alterations in Vibrio fischeri motility correlate with a delay in symbiosis initiation and are associated with additional symbiotic colonization defects. Appl. Environ. Microbiol. 68:2519-2528.
Nishiguchi MK. 2002.Host-symbiont recognition in the environmentally transmitted sepiolid squid-Vibrio mutualism. Microb. Ecol. 44:10-8.
Ackersberg, F., C. Lupp, B. Feliciano and E. G. Ruby. 2001. Vibrio fischeri outer membrane protein OmpU plays a role in normal symbiotic colonization. J. Bacteriol. 183:6590-6597.
Stabb, E. V., K. A. Reich and E. G. Ruby. 2001. Vibrio fischeri genes hvnA and hvnB encode secreted NAD+-Glycohydrolyases. J. Bacteriol. 183:309-317.
Visick, K. L. and L. M. Skoufos. 2001. Two-component sensor required for normal symbiotic colonization of Euprymna scolopes by Vibrio fischeri. J. Bacteriol. 183:835-842.
Callahan, S. M. and P. V. Dunlap. 2000. LuxR- and acyl- homoserine- lactone- controlled non- lux genes define a quorum-sensing regulon in Vibrio fischeri. J. Bacteriol. 182:2811-2822.
Clays, M. F. and P. V. Dunlap. 2000. Aposymbiotic culture of the sepiolid squid Euprymna scolopes: role of symbiotic bacterium Vibrio fischeri in host growth, development, and light organ morphogenesis. J. Exp. Zool. 286:280-296.
Foster, J. S., M. A. Apicella, and M. J. McFall-Ngai. 2000. Vibrio fischeri lipopolysaccharide induces developmental apoptosis, but not complete morphogenesis, of the Euprymna scolopes symbiotic light organ. Dev. Biol. 226:242-254.
Graf, J. and E. G. Ruby. 2000. Novel effects of a transposon insertion in the Vibrio fischeri glnD gene: defects in iron uptake and symbiotic persistence in addition to nitrogen utilization. Mol. Microbiol. 37:168-179.
Lemus, J. D. and M. J. McFall-Ngai. 2000. Alterations in the proteome of the Euprymna scolopes light organ in response to symbiotic Vibrio fischeri.Appl. Environ. Microbiol. 66(9): 4091-4097.
Nyholm, S.V., E. V. Stabb, E. G. Ruby and M. J. McFall-Ngai. 2000. Establishment of an animal-bacterial association: Recruiting symbiotic vibrios from the environment. PNAS 97(18):10231-10235.
Visick, K. L., J. Foster, J. Doino, M. McFall-Ngai and E. G. Ruby. 2000. Vibrio fischeri lux genes play an important role in colonization and development of the host light organ. J. Bacteriol. 182:4578-4586.
Small, A. L. and M. J. McFall-Ngai. 1999. Halide peroxidase in tissues that interact with bacteria in the host squid Euprymna scolopes. J. Cell. Biochem. 72:445-457.
Fidopiastis, P., S. von Boletzky, and E.G. Ruby. 1998. A new niche for Vibrio logei, the predominant light organ symbiont of squids of the genus Sepiola. J. Bacteriol. 180:59-64.
LaMarcq, L.H., and M.J. McFall-Ngai. 1998. Induction of a gradual, reversible morphogenesis of its host’s epithelial brush border by Vibrio fischeri. Infect. Immun. 66:777-785.
Nyholm, S. V. and M. J. McFall-Ngai. 1998. Sampling the light-organ microenvironment of Euprymna scolopes: description of a population of host cells in association with the bacterial symbiont Vibrio fischeri. Biol. Bull. 195:89-97.
Visick, K. L., and E. G. Ruby. 1998. The periplasmic, group III catalase of Vibrio fischeri is required for normal symbiotic competence, and is induced both by oxidative stress and by approach to stationary phase. J. Bacteriol. 180: 2087-2092.
Weis, W.M., A.L. Small, and M.J. McFall-Ngai. 1997. A peroxidase related to the mammalian antimicrobial protein myeloperoxidase in the Euprymna-Vibrio mutualism. Proc. Natl. Acad. Sci. USA. 93:13683-13688.
Boettcher, K.J., E.G. Ruby, and M.J. McFall-Ngai. 1996. Bioluminescence in the symbiotic squid Euprymna scolopes is controlled by a daily biological rhythm. J. Comp. Physiol. A. 179:65-73.
Visick, K. L. and E. G. Ruby. 1996. Construction and symbiotic competence of a luxA-deletion mutant of Vibrio fischeri. Gene 175:89-94.
Boettcher, K. J., and E. G. Ruby. 1995. Detection and quantification of Vibrio fischeri autoinducer from symbiotic squid light organs. J. Bacteriol. 177:1053-1058.
Doino, J.A., and M. J. McFall-Ngai. 1995. A transient exposure to symbiosis-competent bacteria induces light-organ morphogenesis in the host squid. Biol. Bull. 189:347-355.
Graf, J., P. V. Dunlap and E. G. Ruby. 1994. Effect of transposon-induced motility mutations on colonization of the host light organ by Vibrio fischeri. J. Bacteriol. 176:6986-6991.
Montgomery, M. K., and M. J. McFall-Ngai. 1994. Bacterial symbionts induce host organ morphogenesis during early postembryonic development of the squid Euprymna scolopes. Development 120:1719-1729.
Ruby, E. G. and L. M. Asato. 1993. Growth and flagellation of Vibrio fischeri during initiation of the sepiolid squid light organ symbiosis. Arch. Microbiol. 1159:160-167.
McFall-Ngai, M. J. and E. G. Ruby. 1991. Symbiont recognition and subsequent morphogenesis as early events in an animal-bacterial mutualism. Science 254:1491-1494.
Boettcher, K. J. and E. G. Ruby. 1990. Depressed light emission by symbiotic Vibrio fischeri of the sepiolid squid Euprymna scolopes. J. Bacteriol. 172:3701-3706.



{kind=link}
{kind=link}


{kind=link}
Leave a Reply