Tracheal cytotoxin (TCT) is a 921 dalton glycopeptide released by Bordetella pertussis, Vibrio fischeri (as a symbiosis chemical), and Neisseria gonorrhoeae (among other peptidoglycan-derived cytotoxins it produces).
- Cundell, DR; Kanthakumar, K; Taylor, GW; Goldman, WE; Flak, T; Cole, PJ; Wilson, R (1994). “Effect of tracheal cytotoxin from Bordetella pertussis on human neutrophil function in vitro”. Infection and Immunity. 62 (2): 639–43. doi:10.1128/iai.62.2.639-643.1994. PMC 186151. PMID 8300220.
- Koropatnick, TA; Engle, JT; Apicella, MA; Stabb, EV; Goldman, WE; McFall-Ngai, MJ (12 November 2004). “Microbial factor-mediated development in a host-bacterial mutualism”. Science. 306 (5699): 1186–8. Bibcode:2004Sci…306.1186K. doi:10.1126/science.1102218. PMID 15539604. S2CID 41603462.
Here, we report that Vibrio fischeri also releases TCT, which acts in synergy with lipopolysaccharide (LPS) to trigger tissue development in its mutualistic symbiosis with the squid Euprymna scolopes
.
- Cloud, KA; Dillard, JP (June 2002). “A lytic transglycosylase of Neisseria gonorrhoeae is involved in peptidoglycan-derived cytotoxin production”. Infection and Immunity. 70 (6): 2752–7. doi:10.1128/IAI.70.6.2752-2757.2002. PMC 127960. PMID 12010959.
A major peptidoglycan fragment released by gonococci is identical to the tracheal cytotoxin of Bordetella pertussis and has been shown to kill ciliated fallopian tube cells in organ culture.
It is a soluble piece of peptidoglycan (PGN) found in the cell wall of all gram-negative bacteria, but only some bacteria species release TCT due to inability to recycle this piece of anhydromuropeptide.
- Mattoo, S.; Cherry, J. D. (2005). “Molecular Pathogenesis, Epidemiology, and Clinical Manifestations of Respiratory Infections Due to Bordetella pertussis and Other Bordetella Subspecies”. Clinical Microbiology Reviews. 18 (2): 326–82. doi:10.1128/CMR.18.2.326-382.2005. PMC 1082800. PMID 15831828.
- Parkhill, Julian; Sebaihia, Mohammed; Preston, Andrew; Murphy, Lee D; Thomson, Nicholas; Harris, David E; Holden, Matthew T G; Churcher, Carol M; et al. (2003). “Comparative analysis of the genome sequences of Bordetella pertussis, Bordetella parapertussis and Bordetella bronchiseptica”. Nature Genetics. 35 (1): 32–40. doi:10.1038/ng1227. PMID 12910271.
History
In 1980, it was discovered that B. pertussis could attach to hamster tracheal epithelial (HTE) cells, and also, that the supernatant from the cultured bacterium could disrupt the cell cycle of uninfected cells.
- Goldman, W.E. and J.B. Baseman (March 1980). “Abstr. Ann. Meet. Am. Soc. Microbiol”. Abstr. Ann. Meet. Am. Soc. Microbiol.[page needed]
This prompted the scientists W. E. Goldman, D. G. Klapper, and J. B. Baseman to isolate and characterize a novel substance from B. pertussis supernatant. The novel disaccharide tetrapeptide that they had purified showed toxicity for HTE cells and tracheal ring cultures. Subsequently, they named the newly sequestered molecule tracheal cytotoxin (TCT).
- Goldman, William E.; Klapper, David G.; Baseman, Joel B. (1982-05-01). “Detection, isolation, and analysis of a released Bordetella pertussis product toxic to cultured tracheal cells”. Infection and Immunity. 36 (2): 782–94. doi:10.1128/iai.36.2.782-794.1982. PMC 351298. PMID 6177637.

Structure
TCT is a soluble piece of peptidoglycan (PGN) found in the cell wall of all gram-negative bacteria.
- Mattoo, S.; Cherry, J. D. (2005). “Molecular Pathogenesis, Epidemiology, and Clinical Manifestations of Respiratory Infections Due to Bordetella pertussis and Other Bordetella Subspecies”. Clinical Microbiology Reviews. 18 (2): 326–82. doi:10.1128/CMR.18.2.326-382.2005. PMC 1082800. PMID 15831828.
Like all PGNs, TCT is composed of a disaccharide and a peptide chain. The IUPAC name for TCT is N-acetylglucosaminyl-1,6-anhydro-N-acetylmuramyl-(L)-alanyl-γ-(D)-glutamyl-mesodiaminopimelyl-(D)-alanine.
- Cookson, BT; Tyler, AN; Goldman, WE (1989). “Primary structure of the peptidoglycan-derived tracheal cytotoxin of Bordetella pertussis”. Biochemistry. 28 (4): 1744–9. doi:10.1021/bi00430a048. PMID 2541765.

It is classified as a DAP (diaminopimelic acid)-type PGN due to the third amino group within the chain being a diaminopimelyl peptide.
Diaminopimelic acid (DAP) is an amino acid, representing an epsilon-carboxy derivative of lysine. DAP is a characteristic of certain cell walls[Brooks, George H.; Geo F. Brooks (2007). Jawetz, Melnick & Adelberg’s medical microbiology. McGraw-Hill Medical. pp. 85. ISBN 978-0-07-147666-9.] of some bacteria. DAP is often found in the peptide linkages of NAM-NAG chains that make up the cell wall of gram-negative bacteria. When provided, they exhibit normal growth. When in deficiency, they still grow but with the inability to make new cell wall peptidoglycan. This is also the attachment point for Braun’s lipoprotein.[Seltmann, Guntram; Holst, Otto (2002). The Bacterial Cell Wall. Berlin: Springer. pp. 81–82. ISBN 3-540-42608-6.] See also Aspartate-semialdehyde dehydrogenase, an enzyme involved in DAP synthesis and Peptidoglycan and Pimelic acid.
The DAP residue is responsible for directly bonding to the D-alanine peptide of another PGN molecule, thus aiding TCT’s attachment within the cell wall.
- Wilson, R; Read, R; Thomas, M; Rutman, A; Harrison, K; Lund, V; Cookson, B; Goldman, W; et al. (1991-01-01). “Effects of Bordetella pertussis infection on human respiratory epithelium in vivo and in vitro”. Infection and Immunity. 59 (1): 337–45. doi:10.1128/iai.59.1.337-345.1991. PMC 257746. PMID 1987048.
The DAP portion of TCT also implies importance in cytopathogenicity as analogs lacking DAP show a significant reduction in toxicity.
- Luker, K. E.; Collier, J. L.; Kolodziej, E. W.; Marshall, G. R.; Goldman, W. E. (1993). “Bordetella pertussis Tracheal Cytotoxin and Other Muramyl Peptides: Distinct Structure-Activity Relationships for Respiratory Epithelial Cytopathology”. Proceedings of the National Academy of Sciences. 90 (6): 2365–2369. Bibcode:1993PNAS…90.2365L. doi:10.1073/pnas.90.6.2365. PMC 46087. PMID 8460147.
Mechanism of pathogenesis
Most Gram-negative bacteria keep TCT within the cell wall by using a PGN-transporter protein known as AmpG. However, B. pertussis is not capable of recycling PGNs via AmpG and thus, TCT escapes into the surrounding environment.
- Jacobs, C; Joris, B; Jamin, M; Klarsov, K; Van Beeumen, J; Mengin-Lecreulx, D; Van Heijenoort, J; Park, JT; et al. (1995). “AmpD, essential for both beta-lactamase regulation and cell wall recycling, is a novel cytosolic N-acetylmuramyl-L-alanine amidase”. Molecular Microbiology. 15 (3): 553–9. doi:10.1111/j.1365-2958.1995.tb02268.x. PMID 7783625. S2CID 40169153.
- Parkhill, Julian; Sebaihia, Mohammed; Preston, Andrew; Murphy, Lee D; Thomson, Nicholas; Harris, David E; Holden, Matthew T G; Churcher, Carol M; et al. (2003). “Comparative analysis of the genome sequences of Bordetella pertussis, Bordetella parapertussis and Bordetella

Also, TCT appears to be constitutively expressed[dubious – discuss] by B. pertussis.
- Mattoo, S.; Cherry, J. D. (2005). “Molecular Pathogenesis, Epidemiology, and Clinical Manifestations of Respiratory Infections Due to Bordetella pertussis and Other Bordetella Subspecies”. Clinical Microbiology Reviews. 18 (2): 326–82. doi:10.1128/CMR.18.2.326-382.2005. PMC 1082800. PMID 15831828.
The first murine-model studies using TCT involved treatment of hamster tracheal cells. These experiments alluded to TCT’s role in ciliostasis and cellular extrusion of ciliated hamster cells. Also, HTE cells had a markedly reduced level of DNA synthesis post-treatment with TCT.
While previous studies using murine models reported evidence of TCT causing ciliostasis, in vitro studies using human tracheal cells have shown that TCT does not affect ciliary beat frequency of living cells, but instead causes damage and eventual extrusion of ciliated cells.
- Jacobs, C; Joris, B; Jamin, M; Klarsov, K; Van Beeumen, J; Mengin-Lecreulx, D; Van Heijenoort, J; Park, JT; et al. (1995). “AmpD, essential for both beta-lactamase regulation and cell wall recycling, is a novel cytosolic N-acetylmuramyl-L-alanine amidase”. Molecular Microbiology. 15 (3): 553–9. doi:10.1111/j.1365-2958.1995.tb02268.x. PMID 7783625. S2CID 40169153.
In gonorrhea infections, vaginal ciliated epithelial cells have also displayed the same cytopathogenic effects due to TCT recognition.
- Swaminathan, C. P.; Brown, P. H.; Roychowdhury, A.; Wang, Q.; Guan, R.; Silverman, N.; Goldman, W. E.; Boons, G.-J.; Mariuzza, R. A. (2006). “Dual strategies for peptidoglycan discrimination by peptidoglycan recognition proteins (PGRPs)”. Proceedings of the National Academy of Sciences. 103 (3): 684–9. Bibcode:2006PNAS..103..684S. doi:10.1073/pnas.0507656103. PMC 1334652. PMID 16407132.
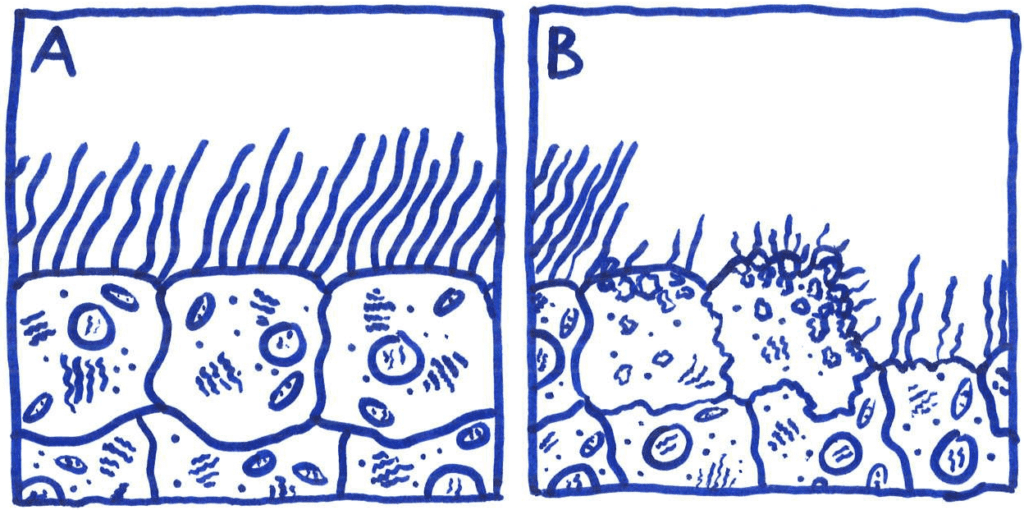
The extensive damage to ciliated epithelial tissue caused by TCT results in major disruption to the ciliary escalator; an important asset of the host’s non-specific defenses. This disruption hinders the host’s ability to remove mucous and foreign microbes from the epithelial tissue. Paroxysmal cough, e.g. whooping cough, is a direct symptom of said mucous build-up due to ciliated tissue damage.
NOD-1 recognition and the presence of Lipooligosaccharide (LOS) are two factors that modulate the effect of TCT. NOD-1 is a pattern recognition receptor that detects peptidoglycan. This receptor reacts weakly to TCT in humans, but robustly in mice. TCT is thought to work synergistically with LOS to mediate an inflammatory response, thus causing damage to ciliated epithelial cells.
- Melvin, Jeffrey A.; Scheller, Erich V.; Miller, Jeff F.; Cotter, Peggy A. (2014-04-01). “Bordetella pertussis pathogenesis: current and future challenges”. Nature Reviews Microbiology. 12 (4): 274–288. doi:10.1038/nrmicro3235. ISSN 1740-1526. PMC 4205565. PMID 24608338.
Notably, the human pathogens (B. pertussis and N. gonorrhea) that produce excess TCT, causing damage to cilia also both produce LOS in their outer membrane.
Lipopolysaccharides are composed of three parts: The O antigen (or O polysaccharide), the Core oligosaccharide, and Lipid A. O-antigen The repetitive glycan polymer contained within an LPS is referred to as the O antigen, O polysaccharide, or O side-chain of the bacteria. The O antigen is attached to the core oligosaccharide, and comprises the outermost domain of the LPS molecule. The composition of the O chain varies from strain to strain; there are over 160 different O antigen structures produced by different E. coli strains.[Raetz CR, Whitfield C (2002). “Lipopolysaccharide endotoxins”. Annu. Rev. Biochem. 71: 635–700. doi:10.1146/annurev.biochem.71.110601.135414. PMC 2569852. PMID 12045108.] The presence or absence of O chains determines whether the LPS is considered “rough” or “smooth”. Full-length O-chains would render the LPS smooth, whereas the absence or reduction of O-chains would make the LPS rough.[ Rittig MG, Kaufmann A, Robins A, Shaw B, Sprenger H, Gemsa D, Foulongne V, Rouot B, Dornand J (2003). “Smooth and rough lipopolysaccharide phenotypes of Brucella induce different intracellular trafficking and cytokine/chemokine release in human monocytes”. J. Leukoc. Biol. 74 (6): 1045–55. doi:10.1189/jlb.0103015. PMID12960272] Bacteria with rough LPS usually have more penetrable cell membranes to hydrophobic antibiotics, since a rough LPS is more hydrophobic.[Tsujimoto H, Gotoh N, Nishino T (1999). “Diffusion of macrolide antibiotics through the outer membrane of Moraxella catarrhalis”. J. Infect. Chemother. 5 (4): 196–200. doi:10.1007/s101560050034. PMID 11810516. S2CID 2742306.] O antigen is exposed on the very outer surface of the bacterial cell, and, as a consequence, is a target for recognition by host antibodies. Core Main article: Core oligosaccharide The core domain always contains an oligosaccharide component that attaches directly to lipid A and commonly contains sugars such as heptose and 3-Deoxy-D-manno-oct-2-ulosonic acid (also known as KDO, keto-deoxyoctulosonate).[Hershberger C, Binkley SB (1968). “Chemistry and metabolism of 3-deoxy-D-mannooctulosonic acid. I. Stereochemical determination”. J. Biol. Chem. 243 (7): 1578–84. doi:10.1016/S0021-9258(18)93581-7. PMID 4296687.] The LPS cores of many bacteria also contain non-carbohydrate components, such as phosphate, amino acids, and ethanolamine substituents. Lipid A Main article: Lipid A Lipid A is, in normal circumstances, a phosphorylated glucosamine disaccharide decorated with multiple fatty acids. These hydrophobic fatty acid chains anchor the LPS into the bacterial membrane, and the rest of the LPS projects from the cell surface. The lipid A domain is responsible for much of the toxicity of Gram-negative bacteria. When bacterial cells are lysed by the immune system, fragments of membrane containing lipid A are released into the circulation, causing fever, diarrhea, and possible fatal endotoxic shock (also called septic shock). The Lipid A moiety is a very conserved component of the LPS.[Tzeng YL, Datta A, Kolli VK, Carlson RW, Stephens DS (May 2002). “Endotoxin of Neisseria meningitidis composed only of intact lipid A: inactivation of the meningococcal 3-deoxy-D-manno-octulosonic acid transferase”. J. Bacteriol. 184 (9): 2379–88. doi:10.1128/JB.184.9.2379-2388.2002. PMC 134985. PMID 11948150.] However Lipid A structure varies among bacterial species. Lipid A structure largely defines the degree and nature of the overall host immune activation.[Khan, Mohd M.; Ernst, Orna; Sun, Jing; Fraser, Iain D. C.; Ernst, Robert K.; Goodlett, David R.; Nita-Lazar, Aleksandra (24 June 2018). “Mass Spectrometry-based Structural Analysis and Systems Immunoproteomics Strategies for Deciphering the Host Response to Endotoxin”. Journal of Molecular Biology. 430 (17): 2641–2660. doi:10.1016/j.jmb.2018.06.032. ISSN 1089-8638. PMID 29949751. S2CID 49481716.] Lipooligosaccharides The “rough form” of Lipopolysaccharides (LPS) has a lower molecular weight due to the absence of the O polysaccharide. In its place is a short oligosaccharide: this form is known as Lipooligosaccharide (LOS), and is a glycolipid found in the outer membrane of some types of Gram-negative bacteria, such as Neisseria spp. and Haemophilus spp.[Moran AP, Prendergast MM, Appelmelk BJ (1996). “Molecular mimicry of host structures by bacterial lipopolysaccharides and its contribution to disease”. FEMS Immunol. Med. Microbiol. 16 (2): 105–15. doi:10.1016/s0928-8244(96)00072-7. PMID 8988391.][Kilár A, Dörnyei Á, Kocsis B (2013). “Structural characterization of bacterial lipopolysaccharides with mass spectrometry and on- and off-line separation techniques”. Mass Spectrom Rev. 32 (2): 90–117. Bibcode:2013MSRv…32…90K. doi:10.1002/mas.21352. PMID 23165926.] LOS plays a central role in maintaining the integrity and functionality of the outer membrane of the Gram negative cell envelope. LOS play an important role in the pathogenesis of certain bacterial infections because they are capable of acting as immunostimulators and immunomodulators.[Moran AP, Prendergast MM, Appelmelk BJ (1996). “Molecular mimicry of host structures by bacterial lipopolysaccharides and its contribution to disease”. FEMS Immunol. Med. Microbiol. 16 (2): 105–15. doi:10.1016/s0928-8244(96)00072-7. PMID 8988391.] Furthermore, LOS molecules are responsible for the ability of some bacterial strains to display molecular mimicry and antigenic diversity, aiding in the evasion of host immune defenses and thus contributing to the virulence of these bacterial strains. In the case of Neisseria meningitidis, the lipid A portion of the molecule has a symmetrical structure and the inner core is composed of 3-deoxy-D-manno-2-octulosonic acid (KDO) and heptose (Hep) moieties. The outer core oligosaccharide chain varies depending on the bacterial strain.[Moran AP, Prendergast MM, Appelmelk BJ (1996). “Molecular mimicry of host structures by bacterial lipopolysaccharides and its contribution to disease”. FEMS Immunol. Med. Microbiol. 16 (2): 105–15. doi:10.1016/s0928-8244(96)00072-7. PMID 8988391.][Kilár A, Dörnyei Á, Kocsis B (2013). “Structural characterization of bacterial lipopolysaccharides with mass spectrometry and on- and off-line separation techniques”. Mass Spectrom Rev. 32 (2): 90–117. Bibcode:2013MSRv…32…90K. doi:10.1002/mas.21352. PMID 23165926.]…Portions of the LPS from several bacterial strains have been shown to be chemically similar to human host cell surface molecules; the ability of some bacteria to present molecules on their surface which are chemically identical or similar to the surface molecules of some types of host cells is termed molecular mimicry.[Chastain EM, Miller SD (2012). “Molecular mimicry as an inducing trigger for CNS autoimmune demyelinating disease”. Immunol. Rev. 245 (1): 227–38. doi:10.1111/j.1600-065X.2011.01076.x. PMC 3586283. PMID 22168423.] For example, in Neisseria meningitidis L2,3,5,7,9, the terminal tetrasaccharide portion of the oligosaccharide (lacto-N-neotetraose) is the same tetrasaccharide as that found in paragloboside, a precursor for ABH glycolipid antigens found on human erythrocytes.[Moran AP, Prendergast MM, Appelmelk BJ (1996). “Molecular mimicry of host structures by bacterial lipopolysaccharides and its contribution to disease”. FEMS Immunol. Med. Microbiol. 16 (2): 105–15. doi:10.1016/s0928-8244(96)00072-7. PMID 8988391.] In another example, the terminal trisaccharide portion (lactotriaose) of the oligosaccharide from pathogenic Neisseria spp. LOS is also found in lactoneoseries glycosphingolipids from human cells.[Moran AP, Prendergast MM, Appelmelk BJ (1996). “Molecular mimicry of host structures by bacterial lipopolysaccharides and its contribution to disease”. FEMS Immunol. Med. Microbiol. 16 (2): 105–15. doi:10.1016/s0928-8244(96)00072-7. PMID 8988391.] Most meningococci from groups B and C, as well as gonococci, have been shown to have this trisaccharide as part of their LOS structure.[Moran AP, Prendergast MM, Appelmelk BJ (1996). “Molecular mimicry of host structures by bacterial lipopolysaccharides and its contribution to disease”. FEMS Immunol. Med. Microbiol. 16 (2): 105–15. doi:10.1016/s0928-8244(96)00072-7. PMID 8988391.] The presence of these human cell surface ‘mimics’ may, in addition to acting as a ‘camouflage’ from the immune system, play a role in the abolishment of immune tolerance when infecting hosts with certain human leukocyte antigen (HLA) genotypes, such as HLA-B35.[Moran AP, Prendergast MM, Appelmelk BJ (1996). “Molecular mimicry of host structures by bacterial lipopolysaccharides and its contribution to disease”. FEMS Immunol. Med. Microbiol. 16 (2): 105–15. doi:10.1016/s0928-8244(96)00072-7. PMID 8988391.] LPS can be sensed directly by hematopoietic stem cells (HSCs) through the bonding with TLR4, causing them to proliferate in reaction to a systemic infection. This response activate the TLR4-TRIF-ROS-p38 signaling within the HSCs and through a sustained TLR4 activation can cause a proliferative stress, leading to impair their competitive repopulating ability.[Takizawa, Hitoshi; Fritsch, Kristin; Kovtonyuk, Larisa V.; Saito, Yasuyuki; Yakkala, Chakradhar; Jacobs, Kurt; Ahuja, Akshay K.; Lopes, Massimo; Hausmann, Annika (3 August 2017). “Pathogen-Induced TLR4-TRIF Innate Immune Signaling in Hematopoietic Stem Cells Promotes Proliferation but Reduces Competitive Fitness”. Cell Stem Cell. 21 (2): 225–240.e5. doi:10.1016/j.stem.2017.06.013. ISSN 1875-9777. PMID 28736216] Infection in mice using S. typhimurium showed similar results, validating the experimental model also in vivo…Normal human blood serum contains anti-LOS antibodies that are bactericidal and patients that have infections caused by serotypically distinct strains possess anti-LOS antibodies that differ in their specificity compared with normal serum.[Yamasaki R, Kerwood DE, Schneider H, Quinn KP, Griffiss JM, Mandrell RE (1994). “The structure of lipooligosaccharide produced by Neisseria gonorrhoeae, strain 15253, isolated from a patient with disseminated infection. Evidence for a new glycosylation pathway of the gonococcal lipooligosaccharide”. J. Biol. Chem. 269 (48): 30345–51. doi:10.1016/S0021-9258(18)43819-7. PMID 7982947.] These differences in humoral immune response to different LOS types can be attributed to the structure of the LOS molecule, primarily within the structure of the oligosaccharide portion of the LOS molecule.[Yamasaki R, Kerwood DE, Schneider H, Quinn KP, Griffiss JM, Mandrell RE (1994). “The structure of lipooligosaccharide produced by Neisseria gonorrhoeae, strain 15253, isolated from a patient with disseminated infection. Evidence for a new glycosylation pathway of the gonococcal lipooligosaccharide”. J. Biol. Chem. 269 (48): 30345–51. doi:10.1016/S0021-9258(18)43819-7. PMID 7982947.] In Neisseria gonorrhoeae it has been demonstrated that the antigenicity of LOS molecules can change during an infection due to the ability of these bacteria to synthesize more than one type of LOS,[Yamasaki R, Kerwood DE, Schneider H, Quinn KP, Griffiss JM, Mandrell RE (1994). “The structure of lipooligosaccharide produced by Neisseria gonorrhoeae, strain 15253, isolated from a patient with disseminated infection. Evidence for a new glycosylation pathway of the gonococcal lipooligosaccharide”. J. Biol. Chem. 269 (48): 30345–51. doi:10.1016/S0021-9258(18)43819-7. PMID 7982947.] a characteristic known as phase variation. Additionally, Neisseria gonorrhoeae, as well as Neisseria meningitidis and Haemophilus influenzae,[Moran AP, Prendergast MM, Appelmelk BJ (1996). “Molecular mimicry of host structures by bacterial lipopolysaccharides and its contribution to disease”. FEMS Immunol. Med. Microbiol. 16 (2): 105–15. doi:10.1016/s0928-8244(96)00072-7. PMID 8988391.] are capable of further modifying their LOS in vitro, for example through sialylation (modification with sialic acid residues), and as a result are able to increase their resistance to complement-mediated killing [Yamasaki R, Kerwood DE, Schneider H, Quinn KP, Griffiss JM, Mandrell RE (1994). “The structure of lipooligosaccharide produced by Neisseria gonorrhoeae, strain 15253, isolated from a patient with disseminated infection. Evidence for a new glycosylation pathway of the gonococcal lipooligosaccharide”. J. Biol. Chem. 269 (48): 30345–51. doi:10.1016/S0021-9258(18)43819-7. PMID 7982947.] or even down-regulate complement activation[Moran AP, Prendergast MM, Appelmelk BJ (1996). “Molecular mimicry of host structures by bacterial lipopolysaccharides and its contribution to disease”. FEMS Immunol. Med. Microbiol. 16 (2): 105–15. doi:10.1016/s0928-8244(96)00072-7. PMID 8988391.] or evade the effects of bactericidal antibodies.[Moran AP, Prendergast MM, Appelmelk BJ (1996). “Molecular mimicry of host structures by bacterial lipopolysaccharides and its contribution to disease”. FEMS Immunol. Med. Microbiol. 16 (2): 105–15. doi:10.1016/s0928-8244(96)00072-7. PMID 8988391.] Sialylation may also contribute to hindered neutrophil attachment and phagocytosis by immune system cells as well as a reduced oxidative burst.[Moran AP, Prendergast MM, Appelmelk BJ (1996). “Molecular mimicry of host structures by bacterial lipopolysaccharides and its contribution to disease”. FEMS Immunol. Med. Microbiol. 16 (2): 105–15. doi:10.1016/s0928-8244(96)00072-7. PMID 8988391.] Haemophilus somnus, a pathogen of cattle, has also been shown to display LOS phase variation, a characteristic which may help in the evasion of bovine host immune defenses.[Howard MD, Cox AD, Weiser JN, Schurig GG, Inzana TJ (2000). “Antigenic diversity of Haemophilus somnus lipooligosaccharide: phase-variable accessibility of the phosphorylcholine epitope”. J. Clin. Microbiol. 38 (12): 4412–9. doi:10.1128/JCM.38.12.4412-4419.2000. PMC 87614. PMID 11101573.] Taken together, these observations suggest that variations in bacterial surface molecules such as LOS can help the pathogen evade both the humoral (antibody and complement-mediated) and the cell-mediated (killing by neutrophils, for example) host immune defenses…In general the health effects of LPS are due to its abilities as a potent activator and modulator of the immune system, especially its inducement of inflammation. Endotoxemia The presence of endotoxins in the blood is called endotoxemia. High level of endotoxemia can lead to septic shock,[Opal SM (2010). “Endotoxins and other sepsis triggers”. Endotoxemia and Endotoxin Shock. Contrib Nephrol. Contributions to Nephrology. Vol. 167. pp. 14–24. doi:10.1159/000315915. ISBN 978-3-8055-9484-4. PMID 20519895.] while lower concentration of endotoxins in the bloodstream is called metabolic endotoxemia.[Gomes, Júnia Maria Geraldo; Costa, Jorge de Assis; Alfenas, Rita de Cássia Gonçalves (March 2017). “Metabolic endotoxemia and diabetes mellitus: A systematic review”. Metabolism. 68: 133–144. doi:10.1016/j.metabol.2016.12.009. PMID 28183445.] Endotoxemia is associated with obesity, diet,[Kallio, K. A. Elisa; Hätönen, Katja A.; Lehto, Markku; Salomaa, Veikko; Männistö, Satu; Pussinen, Pirkko J. (April 2015). “Endotoxemia, nutrition, and cardiometabolic disorders”. Acta Diabetologica. 52 (2): 395–404. doi:10.1007/s00592-014-0662-3. ISSN 0940-5429. PMID 25326898. S2CID 24020127.] cardiovascular diseases,[Kallio, K. A. Elisa; Hätönen, Katja A.; Lehto, Markku; Salomaa, Veikko; Männistö, Satu; Pussinen, Pirkko J. (April 2015). “Endotoxemia, nutrition, and cardiometabolic disorders”. Acta Diabetologica. 52 (2): 395–404. doi:10.1007/s00592-014-0662-3. ISSN 0940-5429. PMID 25326898. S2CID 24020127.] and diabetes,[Gomes, Júnia Maria Geraldo; Costa, Jorge de Assis; Alfenas, Rita de Cássia Gonçalves (March 2017). “Metabolic endotoxemia and diabetes mellitus: A systematic review”. Metabolism. 68: 133–144. doi:10.1016/j.metabol.2016.12.009. PMID 28183445.] while also host genetics might have an effect.[Leskelä, Jaakko; Toppila, Iiro; Härma, Mari‐Anne; Palviainen, Teemu; Salminen, Aino; Sandholm, Niina; Pietiäinen, Milla; Kopra, Elisa; Pais de Barros, Jean‐Paul; FinnGen; Lassenius, Mariann I. (20 October 2021). “Genetic Profile of Endotoxemia Reveals an Association With Thromboembolism and Stroke”. Journal of the American Heart Association. 10 (21): e022482. doi:10.1161/JAHA.121.022482. ISSN 2047-9980. PMC 8751832. PMID 34668383.] Moreover, endotoxemia of intestinal origin, especially, at the host-pathogen interface, is considered to be an important factor in the development of alcoholic hepatitis,[Ceccanti M, Attili A, Balducci G, Attilia F, Giacomelli S, Rotondo C, Sasso GF, Xirouchakis E, Attilia ML (2006). “Acute alcoholic hepatitis”. J. Clin. Gastroenterol. 40 (9): 833–41. doi:10.1097/01.mcg.0000225570.04773.5d. PMID 17016141.] which is likely to develop on the basis of the small bowel bacterial overgrowth syndrome and an increased intestinal permeability.[Parlesak A, Schäfer C, Schütz T, Bode JC, Bode C (2000). “Increased intestinal permeability to macromolecules and endotoxemia in patients with chronic alcohol abuse in different stages of alcohol-induced liver disease”. J. Hepatol. 32 (5): 742–7. doi:10.1016/S0168-8278(00)80242-1. PMID 10845660.] Lipid A may cause uncontrolled activation of mammalian immune systems with production of inflammatory mediators that may lead to septic shock.[Kilár A, Dörnyei Á, Kocsis B (2013). “Structural characterization of bacterial lipopolysaccharides with mass spectrometry and on- and off-line separation techniques”. Mass Spectrom Rev. 32 (2): 90–117. Bibcode:2013MSRv…32…90K. doi:10.1002/mas.21352. PMID 23165926.] This inflammatory reaction is mediated by Toll-like receptor 4 which is responsible for immune system cell activation.[Kilár A, Dörnyei Á, Kocsis B (2013). “Structural characterization of bacterial lipopolysaccharides with mass spectrometry and on- and off-line separation techniques”. Mass Spectrom Rev. 32 (2): 90–117. Bibcode:2013MSRv…32…90K. doi:10.1002/mas.21352. PMID 23165926.] Damage to the endothelial layer of blood vessels caused by these inflammatory mediators can lead to capillary leak syndrome, dilation of blood vessels and a decrease in cardiac function and can lead to septic shock.[Stephens DS, Greenwood B, Brandtzaeg P (2007). “Epidemic meningitis, meningococcaemia, and Neisseria meningitidis”. Lancet. 369 (9580): 2196–210. doi:10.1016/S0140-6736(07)61016-2. PMID 17604802. S2CID 16951072.] Pronounced complement activation can also be observed later in the course as the bacteria multiply in the blood.[Stephens DS, Greenwood B, Brandtzaeg P (2007). “Epidemic meningitis, meningococcaemia, and Neisseria meningitidis”. Lancet. 369 (9580): 2196–210. doi:10.1016/S0140-6736(07)61016-2. PMID 17604802. S2CID 16951072.] High bacterial proliferation triggering destructive endothelial damage can also lead to disseminated intravascular coagulation (DIC) with loss of function of certain internal organs such as the kidneys, adrenal glands and lungs due to compromised blood supply. The skin can show the effects of vascular damage often coupled with depletion of coagulation factors in the form of petechiae, purpura and ecchymoses. The limbs can also be affected, sometimes with devastating consequences such as the development of gangrene, requiring subsequent amputation.[Stephens DS, Greenwood B, Brandtzaeg P (2007). “Epidemic meningitis, meningococcaemia, and Neisseria meningitidis”. Lancet. 369 (9580): 2196–210. doi:10.1016/S0140-6736(07)61016-2. PMID 17604802. S2CID 16951072.] Loss of function of the adrenal glands can cause adrenal insufficiency and additional hemorrhage into the adrenals causes Waterhouse-Friderichsen syndrome, both of which can be life-threatening. It has also been reported that gonococcal LOS can cause damage to human fallopian tubes.[Yamasaki R, Kerwood DE, Schneider H, Quinn KP, Griffiss JM, Mandrell RE (1994). “The structure of lipooligosaccharide produced by Neisseria gonorrhoeae, strain 15253, isolated from a patient with disseminated infection. Evidence for a new glycosylation pathway of the gonococcal lipooligosaccharide”. J. Biol. Chem. 269 (48): 30345–51. doi:10.1016/S0021-9258(18)43819-7. PMID 7982947.] Auto-immune disease The molecular mimicry of some LOS molecules is thought to cause autoimmune-based host responses, such as flareups of multiple sclerosis.[Moran AP, Prendergast MM, Appelmelk BJ (1996). “Molecular mimicry of host structures by bacterial lipopolysaccharides and its contribution to disease”. FEMS Immunol. Med. Microbiol. 16 (2): 105–15. doi:10.1016/s0928-8244(96)00072-7. PMID 8988391.][Chastain EM, Miller SD (2012). “Molecular mimicry as an inducing trigger for CNS autoimmune demyelinating disease”. Immunol. Rev. 245 (1): 227–38. doi:10.1111/j.1600-065X.2011.01076.x. PMC 3586283. PMID 22168423.] Other examples of bacterial mimicry of host structures via LOS are found with the bacteria Helicobacter pylori and Campylobacter jejuni, organisms which cause gastrointestinal disease in humans, and Haemophilus ducreyi which causes chancroid. Certain C. jejuni LPS serotypes (attributed to certain tetra- and pentasaccharide moieties of the core oligosaccharide) have also been implicated with Guillain–Barré syndrome and a variant of Guillain–Barré called Miller-Fisher syndrome.[Moran AP, Prendergast MM, Appelmelk BJ (1996). “Molecular mimicry of host structures by bacterial lipopolysaccharides and its contribution to disease”. FEMS Immunol. Med. Microbiol. 16 (2): 105–15. doi:10.1016/s0928-8244(96)00072-7. PMID 8988391.] Link to obesity Epidemiological studies have shown that increased endotoxin load, which can be a result of increased populations of endotoxin-producing bacteria in the intestinal tract, is associated with certain obesity-related patient groups.[Moreno-Navarrete JM, Ortega F, Serino M, Luche E, Waget A, Pardo G, Salvador J, Ricart W, Frühbeck G, Burcelin R, Fernández-Real JM (2012). “Circulating lipopolysaccharide-binding protein (LBP) as a marker of obesity-related insulin resistance”. Int J Obes (Lond). 36 (11): 1442–9. doi:10.1038/ijo.2011.256. PMID 22184060.][Lepper PM, Schumann C, Triantafilou K, Rasche FM, Schuster T, Frank H, Schneider EM, Triantafilou M, von Eynatten M (2007). “Association of lipopolysaccharide-binding protein and coronary artery disease in men”. J. Am. Coll. Cardiol. 50 (1): 25–31. doi:10.1016/j.jacc.2007.02.070. PMID 17601541.][Ruiz AG, Casafont F, Crespo J, Cayón A, Mayorga M, Estebanez A, Fernadez-Escalante JC, Pons-Romero F (2007). “Lipopolysaccharide-binding protein plasma levels and liver TNF-alpha gene expression in obese patients: evidence for the potential role of endotoxin in the pathogenesis of non-alcoholic steatohepatitis”. Obes Surg. 17 (10): 1374–80. doi:10.1007/s11695-007-9243-7. PMID 18000721. S2CID 44494003.] Other studies have shown that purified endotoxin from Escherichia coli can induce obesity and insulin-resistance when injected into germ-free mouse models.[Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, Neyrinck AM, Fava F, Tuohy KM, Chabo C, Waget A, Delmée E, Cousin B, Sulpice T, Chamontin B, Ferrières J, Tanti JF, Gibson GR, Casteilla L, Delzenne NM, Alessi MC, Burcelin R (2007). “Metabolic endotoxemia initiates obesity and insulin resistance”. Diabetes. 56 (7): 1761–72. doi:10.2337/db06-1491. PMID 17456850.] A more recent study has uncovered a potentially contributing role for Enterobacter cloacae B29 toward obesity and insulin resistance in a human patient.[Fei N, Zhao L (December 2012). “An opportunistic pathogen isolated from the gut of an obese human causes obesity in germfree mice”. ISME J. 7 (4): 880–4. doi:10.1038/ismej.2012.153. PMC 3603399. PMID 23235292.] The presumed mechanism for the association of endotoxin with obesity is that endotoxin induces an inflammation-mediated pathway accounting for the observed obesity and insulin resistance.[Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, Neyrinck AM, Fava F, Tuohy KM, Chabo C, Waget A, Delmée E, Cousin B, Sulpice T, Chamontin B, Ferrières J, Tanti JF, Gibson GR, Casteilla L, Delzenne NM, Alessi MC, Burcelin R (2007). “Metabolic endotoxemia initiates obesity and insulin resistance”. Diabetes. 56 (7): 1761–72. doi:10.2337/db06-1491. PMID 17456850.] Bacterial genera associated with endotoxin-related obesity effects include Escherichia and Enterobacter. Depression There is experimental and observational evidence that LPS might play a role in depression. Administration of LPS in mice can lead to depressive symptoms, and there seem to be elevated levels of LPS in some people with depression. Inflammation may sometimes play a role in the development of depression, and LPS is pro-inflammatory.[Lasselin, Julie; Schedlowski, Manfred; Karshikoff, Bianka; Engler, Harald; Lekander, Mats; Konsman, Jan Pieter (August 2020). “Comparison of bacterial lipopolysaccharide-induced sickness behavior in rodents and humans: Relevance for symptoms of anxiety and depression”. Neuroscience and Biobehavioral Reviews. 115: 15–24. doi:10.1016/j.neubiorev.2020.05.001. ISSN 1873-7528. PMID 32433924. S2CID 218665128.] Cellular senescence Inflammation induced by LPS can induce cellular senescence, as has been shown for the lung epithelial cells and microglial cells (the latter leading to neurodegeneration).[Wei W, Ji S (2018). “Cellular senescence: Molecular mechanisms and pathogenicity”. Journal of Cellular Physiology. 233 (12): 9121–9135. doi:10.1002/jcp.26956. PMID 30078211. S2CID 51924586.] Role as contaminant in biotechnology and research Lipopolysaccharides are frequent contaminants in plasmid DNA prepared from bacteria or proteins expressed from bacteria, and must be removed from the DNA or protein to avoid contaminating experiments and to avoid toxicity of products manufactured using industrial fermentation.[Wicks, Ian P.; Howell, Meredith L.; Hancock, Tuesday; Kohsaka, Hitoshi; Olee, Tsaiwei; Carson, Dennis A. (March 1995). “Bacterial Lipopolysaccharide Copurifies with Plasmid DNA: Implications for Animal Models and Human Gene Therapy”. Human Gene Therapy. 6 (3): 317–323. doi:10.1089/hum.1995.6.3-317. PMID 7779915.] Ovalbumin is frequently contaminated with endotoxins. Ovalbumin is one of the extensively studied proteins in animal models and also an established model allergen for airway hyper-responsiveness (AHR). Commercially available ovalbumin that is contaminated with LPS can falsify research results, as it does not accurately reflect the effect of the protein antigen on animal physiology.[Watanabe, Junji; Miyazaki, Yasunari; Zimmerman, Guy A.; Albertine, Kurt H.; McIntyre, Thomas M. (24 October 2003). “Endotoxin contamination of ovalbumin suppresses murine immunologic responses and development of airway hyper-reactivity”. The Journal of Biological Chemistry. 278 (43): 42361–42368. doi:10.1074/jbc.M307752200. ISSN 0021-9258. PMID 12909619.] In pharmaceutical production, it is necessary to remove all traces of endotoxin from drug product containers, as even small amounts of endotoxin will cause illness in humans. A depyrogenation oven is used for this purpose. Temperatures in excess of 300 °C are required to fully break down LPS.[16 December 2014. “The Detection of Endotoxins Via the LAL Test, the Chromogenic Method”. Archived from the original on 29 March 2015. Retrieved 14 March 2015.] The standard assay for detecting presence of endotoxin is the Limulus Amebocyte Lysate (LAL) assay, utilizing blood from the Horseshoe crab (Limulus polyphemus).[Iwanaga S (2007). “Biochemical principle of Limulus test for detecting bacterial endotoxins”. Proc. Jpn. Acad., Ser. B, Phys. Biol. Sci. 83 (4): 110–9. Bibcode:2007PJAB…83..110I. doi:10.2183/pjab.83.110. PMC 3756735. PMID 24019589.]
- Limulus amebocyte lysate (LAL) is an aqueous extract of blood cells (amoebocytes) from the Atlantic horseshoe crab Limulus polyphemus. LAL reacts with bacterial endotoxin lipopolysaccharide (LPS), which is a membrane component of gram-negative bacteria. This reaction is the basis of the LAL test, which is widely used for the detection and quantification of bacterial endotoxins.In Asia, a similar Tachypleus amebocyte lysate (TAL) test based on the local horseshoe crabs Tachypleus gigas or Tachypleus tridentatus is occasionally used instead.[Sheng J, Zhou J, Peng Y, Zhu Z, Chen L (1 January 2006). “Tachypleus Amebocyte Lysate Test Using in Transfusion Reaction”. Chinese Journal of Nosocomiology (in Chinese).] The recombinant factor C (rFC) assay is a replacement of LAL/TAL based on a similar reaction.[ “Endotoxin Testing Manufacturers and Conservation”. www.horseshoecrab.org. Retrieved 10 March 2020.] Background American medical researcher Fred Bang reported in 1956 that gram-negative bacteria, even if killed, will cause the blood of the horseshoe crab to turn into a semi-solid mass. It was later recognized that the animal’s blood cells, mobile cells called amoebocytes, contain granules with a clotting factor known as coagulogen; this is released outside the cell when bacterial endotoxin is encountered. The resulting coagulation (gelling) is thought to contain bacterial infections in the animal’s semi-closed circulatory system.[Greer S. “Frederik Bang”. JH Bloomberg School of Public Health. Johns Hopkins Bloomberg School of Public Health.] Modern analysis of the lysate has led to understanding of this system of cascade, with multiple enzymes working in sequence to produce the gel. The entry point of endotoxin-induced clotting is Limulus clotting factor C.[Iwanaga S (May 2007). “Biochemical principle of Limulus test for detecting bacterial endotoxins”. Proceedings of the Japan Academy. Series B, Physical and Biological Sciences. 83 (4): 110–9. Bibcode:2007PJAB…83..110I. doi:10.2183/pjab.83.110. PMC 3756735. PMID 24019589.] In 1977 the U.S. Food and Drug Administration (FDA) approved LAL for testing drugs, products and devices that come in contact with the blood. Prior to that date, a much slower and more expensive test on rabbits had been used for this purpose.[Zhang S (9 May 2018). “The Last Days of the Blue-Blood Harvest”. The Atlantic. Retrieved 15 May 2018.]
- Limulus clotting enzyme (EC 3.4.21.86, clotting enzyme) is an enzyme.[Muta T, Hashimoto R, Miyata T, Nishimura H, Toh Y, Iwanaga S (December 1990). “Proclotting enzyme from horseshoe crab hemocytes. cDNA cloning, disulfide locations, and subcellular localization”. The Journal of Biological Chemistry. 265 (36): 22426–33. doi:10.1016/S0021-9258(18)45722-5. PMID 2266134.][Tokunaga F, Nakajima H, Iwanaga S (January 1991). “Further studies on lipopolysaccharide-sensitive serine protease zymogen (factor C): its isolation from Limulus polyphemus hemocytes and identification as an intracellular zymogen activated by alpha-chymotrypsin, not by trypsin”. Journal of Biochemistry. 109 (1): 150–7. doi:10.1093/oxfordjournals.jbchem.a123337. PMID 2016264.] This enzyme catalyses the following chemical reactionSelective cleavage of -Arg18– and -Arg47– bonds in coagulogen to form coagulin and fragments. This enzyme is present in the hemocyte granules of horseshoe crabs Limulus and Tachypleus. In the immunity-related clotting pathways of these organisms, it is the final enzyme responsible for the activation of coagulin.[Iwanaga, S (May 2007). “Biochemical principle of Limulus test for detecting bacterial endotoxins”. Proceedings of the Japan Academy. Series B, Physical and Biological Sciences. 83 (4): 110–9. Bibcode:2007PJAB…83..110I. doi:10.2183/pjab.83.110. PMC 3756735. PMID 24019589.]
- Coagulin is a gel-forming protein of hemolymph that hinders the spread of invaders by immobilising them. It is produced in the coagulogen form before being cleaved into the active form.[Iwanaga S, Miyata T, Srimal S, Kawabata S (1985). “The complete amino acid sequence of coagulogen isolated from Southeast Asian horseshoe crab, Carcinoscorpius rotundicauda”. J. Biochem. 98 (2): 305–318. doi:10.1093/oxfordjournals.jbchem.a135283. PMID 3905780.][Iwanaga S, Miyata T, Usui K (1984). “The amino acid sequence of coagulogen isolated from southeast Asian horseshoe crab, Tachypleus gigas”. J. Biochem. 95 (6): 1793–1801. doi:10.1093/oxfordjournals.jbchem.a134792. PMID 6469947.] In human medicine, coagulation of coagulin is the basis of detection of bacterial endotoxin in the LAL test for parenteral medications. The protein contains a single 175-residue polypeptide chain that is cleaved after Arg-18 and Arg-46 by a Limulus clotting enzyme contained in the hemocyte and activated by a bacterial endotoxin (lipopolysaccharide). Cleavage releases two chains of coagulin, A and B, linked by two disulfide bonds, together with the peptide C.[Iwanaga S, Miyata T, Srimal S, Kawabata S (1985). “The complete amino acid sequence of coagulogen isolated from Southeast Asian horseshoe crab, Carcinoscorpius rotundicauda”. J. Biochem. 98 (2): 305–318. doi:10.1093/oxfordjournals.jbchem.a135283. PMID 3905780.][Iwanaga S, Miyata T, Usui K (1984). “The amino acid sequence of coagulogen isolated from southeast Asian horseshoe crab, Tachypleus gigas”. J. Biochem. 95 (6): 1793–1801. doi:10.1093/oxfordjournals.jbchem.a134792. PMID 6469947.] Gel formation results from interlinking of coagulin molecules.[Iwanaga S, Miyata T, Srimal S, Kawabata S (1985). “The complete amino acid sequence of coagulogen isolated from Southeast Asian horseshoe crab, Carcinoscorpius rotundicauda”. J. Biochem. 98 (2): 305–318. doi:10.1093/oxfordjournals.jbchem.a135283. PMID 3905780.] The full-length structure of a coagulogen is known (PDB: 1AOC); it shares the same cystine-knot cytokine superfamily (fold) as neurotrophins, with several cystines conserved. The A-B fold wraps around the helical peptide C, forming a compact structure.[Bergner, A; Oganessyan, V; Muta, T; Iwanaga, S; Typke, D; Huber, R; Bode, W (16 December 1996). “Crystal structure of a coagulogen, the clotting protein from horseshoe crab: a structural homologue of nerve growth factor”. The EMBO Journal. 15 (24): 6789–97. doi:10.1002/j.1460-2075.1996.tb01070.x. ISSN 0261-4189. PMC 452504. PMID 9003754.] In crustaceans, hemolymph coagulation depends on the transglutaminase-mediated cross-linking of specific plasma-clotting proteins, but without the proteolytic cascade. In horseshoe crabs, the proteolytic coagulation cascade triggered by lipopolysaccharides and beta-1,3-glucans leads to the conversion of coagulogen into coagulin, resulting in noncovalent coagulin homopolymers through head-to-tail interaction. However, horseshoe crab transglutaminase does not cross-link coagulins intermolecularly. Recently, coagulins were discovered to be cross-linked on hemocyte cell surface proteins called proxins. This indicates that a cross-linking reaction at the final stage of hemolymph coagulation is an important innate immune system of horseshoe crabs.[Kawabata S, Osaki T (2004). “Structure and function of coagulogen, a clottable protein in horseshoe crabs”. Cell. Mol. Life Sci. 61 (11): 1257–1265. doi:10.1007/s00018-004-3396-5. PMID 15170505. S2CID 24537601.] In comparison, mammalian blood coagulation is based on the proteolytically induced polymerization of fibrinogens. Initially, fibrin monomers noncovalently interact with each other. The resulting homopolymers are further stabilized when the plasma transglutaminase cross-links epsilon-(gamma-glutamyl)lysine bonds between molecules.
- Limulus clotting enzyme (EC 3.4.21.86, clotting enzyme) is an enzyme.[Muta T, Hashimoto R, Miyata T, Nishimura H, Toh Y, Iwanaga S (December 1990). “Proclotting enzyme from horseshoe crab hemocytes. cDNA cloning, disulfide locations, and subcellular localization”. The Journal of Biological Chemistry. 265 (36): 22426–33. doi:10.1016/S0021-9258(18)45722-5. PMID 2266134.][Tokunaga F, Nakajima H, Iwanaga S (January 1991). “Further studies on lipopolysaccharide-sensitive serine protease zymogen (factor C): its isolation from Limulus polyphemus hemocytes and identification as an intracellular zymogen activated by alpha-chymotrypsin, not by trypsin”. Journal of Biochemistry. 109 (1): 150–7. doi:10.1093/oxfordjournals.jbchem.a123337. PMID 2016264.] This enzyme catalyses the following chemical reactionSelective cleavage of -Arg18– and -Arg47– bonds in coagulogen to form coagulin and fragments. This enzyme is present in the hemocyte granules of horseshoe crabs Limulus and Tachypleus. In the immunity-related clotting pathways of these organisms, it is the final enzyme responsible for the activation of coagulin.[Iwanaga, S (May 2007). “Biochemical principle of Limulus test for detecting bacterial endotoxins”. Proceedings of the Japan Academy. Series B, Physical and Biological Sciences. 83 (4): 110–9. Bibcode:2007PJAB…83..110I. doi:10.2183/pjab.83.110. PMC 3756735. PMID 24019589.]
Very low levels of LPS can cause coagulation of the limulus lysate due to a powerful amplification through an enzymatic cascade. However, due to the dwindling population of horseshoe crabs, and the fact that there are factors that interfere with the LAL assay, efforts have been made to develop alternative assays, with the most promising ones being ELISA tests using a recombinant version of a protein in the LAL assay, Factor C.[Ding JL, Ho B (2001). “A new era in pyrogen testing” (PDF). Trends Biotechnol. 19 (8): 277–81. doi:10.1016/s0167-7799(01)01694-8. PMID 11451451. Archived from the original (PDF) on 2 January 2014. Retrieved 2 January 2014.] See also Bioaerosol and Depyrogenation and Host-pathogen interface and Mucopolysaccharide and Nesfatin-1 and Schwartzman reaction and AOAH.
Effect on immune system
TCT has been classified as an adjuvant molecule because of the stimulating effects it has on the immune system. Cellular damage associated with TCT is thought to be a result of increased levels of nitric oxide (NO) secretion by mucosal cells as part of an innate defense response to extracellular lipopolysaccharide (LPS) and TCT.
- Coote, JG (2001). Environmental sensing mechanisms in Bordetella. Advances in Microbial Physiology. Vol. 44. pp. 141–81. doi:10.1016/S0065-2911(01)44013-6. ISBN 9780120277445. PMID 11407112.
In humans, peptidoglycan recognition proteins, e.g. PGRPIαC, appear to bind with TCT and consequently induce the Tumor Necrosis Factor Receptor (TNFR) pathway.
- Swaminathan, C. P.; Brown, PH; Roychowdhury, A; Wang, Q; Guan, R; Silverman, N; Goldman, WE; Boons, GJ; Mariuzza, RA (2006). “Dual strategies for peptidoglycan discrimination by peptidoglycan recognition proteins (PGRPs)”. Proceedings of the National Academy of Sciences. 103 (3): 684–9. Bibcode:2006PNAS..103..684S. doi:10.1073/pnas.0507656103. PMC 1334652. PMID 16407132.
Studies using murine macrophages have shown that TCT encourages cytokine secretion, probably through the NOD1 receptor.
- Humann, Jessica; Lenz, Laurel L. (2009). “Bacterial Peptidoglycan-Degrading Enzymes and Their Impact on Host Muropeptide Detection”. Journal of Innate Immunity. 1 (2): 88–97. doi:10.1159/000181181. PMC 2659621. PMID 19319201.
As a pleiotropic toxin, TCT also acts as a pyrogen and as a stimulant of slow-wave sleep.
- Magalhaes, Joao Gamelas; Philpott, Dana J; Nahori, Marie-Anne; Jéhanno, Muguette; Fritz, Joerg; Bourhis, Lionel Le; Viala, Jérôme; Hugot, Jean-Pierre; et al. (2005). “Murine Nod1 but not its human orthologue mediates innate immune detection of tracheal cytotoxin”. EMBO
Pleiotropy (from Greek πλείων pleion, ‘more’, and τρόπος tropos, ‘way’) occurs when one gene influences two or more seemingly unrelated phenotypic traits. Such a gene that exhibits multiple phenotypic expression is called a pleiotropic gene. Mutation in a pleiotropic gene may have an effect on several traits simultaneously, due to the gene coding for a product used by a myriad of cells or different targets that have the same signaling function. Pleiotropy can arise from several distinct but potentially overlapping mechanisms, such as gene pleiotropy, developmental pleiotropy, and selectional pleiotropy. Gene pleiotropy occurs when a gene product interacts with multiple other proteins or catalyzes multiple reactions. Developmental pleiotropy occurs when mutations have multiple effects on the resulting phenotype. Selectional pleiotropy occurs when the resulting phenotype has many effects on fitness (depending on factors such as age and gender).[Paaby, Annalise B.; Rockman, Matthew V. (2016-11-15). “The many faces of pleiotropy”. Trends in Genetics. 29 (2): 66–73. doi:10.1016/j.tig.2012.10.010. PMC 3558540. PMID 23140989.] An example of pleiotropy is phenylketonuria, an inherited disorder that affects the level of phenylalanine, an amino acid that can be obtained from food (this ‘inherited’ disorder can be acquired and has been induced through feeding). [ “Phenylketonuria”. Genes and Disease. National Center for Biotechnology Information. 1998.] That page goes on to discuss albinism, autism, schizophrenia, sickle cell anemia, marfun syndrome, mini-muscle allele, dna repair proteins and chickens In pharmacology, pleiotropy includes all of a drug’s actions other than those for which the agent was specifically developed. It may include adverse effects which are detrimental ones,[Davignon J (June 2004). “Beneficial cardiovascular pleiotropic effects of statins”. Circulation. 109 (23 Suppl 1): III39–43. doi:10.1161/01.CIR.0000131517.20177.5a. PMID 15198965.] but is often used to denote additional beneficial effects.[Rod Flower; Humphrey P. Rang; Maureen M. Dale; Ritter, James M. (2007). Rang & Dale’s pharmacology. Edinburgh: Churchill Livingstone. ISBN 978-0-443-06911-6.] Steroid drugs, such as prednisone and prednisolone, have pleiotropic effects, including systemic ones, for the same reason that endogenous steroid hormones do: cells throughout the body have receptors that can respond to them, because the endogenous ones are endocrine messengers.
Peptidoglycan recognition protein 4 (PGLYRP4), in mammals (mice), interacts with TCT and reduces damage from pertussis inflammation. This molecule has similar immune-eliciting properties in Drosophila, where a pair of PGRPs perform the recognition.
- Skerry, C; Goldman, WE; Carbonetti, NH (February 2019). “Peptidoglycan Recognition Protein 4 Suppresses Early Inflammatory Responses to Bordetella pertussis and Contributes to Sphingosine-1-Phosphate Receptor Agonist-Mediated Disease Attenuation”. Infection and Immunity. 87 (2). doi:10.1128/IAI.00601-18. PMC 6346131. PMID 30510103.
- Chang, CI; Chelliah, Y; Borek, D; Mengin-Lecreulx, D; Deisenhofer, J (24 March 2006). “Structure of tracheal cytotoxin in complex with a heterodimeric pattern-recognition receptor”. Science. 311 (5768): 1761–4. Bibcode:2006Sci…311.1761C. doi:10.1126/science.1123056. PMID 16556841. S2CID 25416989.
PGLYRP4 (formerly PGRP-Iβ), a member of a family of human Peptidoglycan Recognition Proteins (PGRPs), was discovered in 2001 by Roman Dziarski and coworkers who cloned and identified the genes for three human PGRPs, PGRP-L, PGRP-Iα, and PGRP-Iβ (named for long and intermediate size transcripts),[Liu C, Xu Z, Gupta D, Dziarski R (September 2001). “Peptidoglycan recognition proteins: a novel family of four human innate immunity pattern recognition molecules”. The Journal of Biological Chemistry. 276 (37): 34686–94. doi:10.1074/jbc.M105566200. PMID 11461926. S2CID 44619852.] and established that human genome codes for a family of 4 PGRPs: PGRP-S (short PGRP or PGRP-S) [Kang D, Liu G, Lundström A, Gelius E, Steiner H (August 1998). “A peptidoglycan recognition protein in innate immunity conserved from insects to humans”. Proceedings of the National Academy of Sciences of the United States of America. 95 (17): 10078–82. Bibcode:1998PNAS…9510078K. doi:10.1073/pnas.95.17.10078. PMC 21464. PMID 9707603.] and PGRP-L, PGRP-Iα, and PGRP-Iβ.[Liu C, Xu Z, Gupta D, Dziarski R (September 2001). “Peptidoglycan recognition proteins: a novel family of four human innate immunity pattern recognition molecules”. The Journal of Biological Chemistry. 276 (37): 34686–94. doi:10.1074/jbc.M105566200. PMID 11461926. S2CID 44619852.] Subsequently, the Human Genome Organization Gene Nomenclature Committee changed the gene symbols of PGRP-S, PGRP-L, PGRP-Iα, and PGRP-Iβ to PGLYRP1 (peptidoglycan recognition protein 1), PGLYRP2 (peptidoglycan recognition protein 2), PGLYRP3 (peptidoglycan recognition protein 3), and PGLYRP4 (peptidoglycan recognition protein 4), respectively, and this nomenclature is currently also used for other mammalian PGRPs.
References
- Cundell, DR; Kanthakumar, K; Taylor, GW; Goldman, WE; Flak, T; Cole, PJ; Wilson, R (1994). “Effect of tracheal cytotoxin from Bordetella pertussis on human neutrophil function in vitro”. Infection and Immunity. 62 (2): 639–43. doi:10.1128/iai.62.2.639-643.1994. PMC 186151. PMID 8300220.
- Koropatnick, TA; Engle, JT; Apicella, MA; Stabb, EV; Goldman, WE; McFall-Ngai, MJ (12 November 2004). “Microbial factor-mediated development in a host-bacterial mutualism”. Science. 306 (5699): 1186–8. Bibcode:2004Sci…306.1186K. doi:10.1126/science.1102218. PMID 15539604. S2CID 41603462.
Here, we report that Vibrio fischeri also releases TCT, which acts in synergy with lipopolysaccharide (LPS) to trigger tissue development in its mutualistic symbiosis with the squid Euprymna scolopes.
- Cloud, KA; Dillard, JP (June 2002). “A lytic transglycosylase of Neisseria gonorrhoeae is involved in peptidoglycan-derived cytotoxin production”. Infection and Immunity. 70 (6): 2752–7. doi:10.1128/IAI.70.6.2752-2757.2002. PMC 127960. PMID 12010959.
A major peptidoglycan fragment released by gonococci is identical to the tracheal cytotoxin of Bordetella pertussis and has been shown to kill ciliated fallopian tube cells in organ culture.
- Mattoo, S.; Cherry, J. D. (2005). “Molecular Pathogenesis, Epidemiology, and Clinical Manifestations of Respiratory Infections Due to Bordetella pertussis and Other Bordetella Subspecies”. Clinical Microbiology Reviews. 18 (2): 326–82. doi:10.1128/CMR.18.2.326-382.2005. PMC 1082800. PMID 15831828.
- Parkhill, Julian; Sebaihia, Mohammed; Preston, Andrew; Murphy, Lee D; Thomson, Nicholas; Harris, David E; Holden, Matthew T G; Churcher, Carol M; et al. (2003). “Comparative analysis of the genome sequences of Bordetella pertussis, Bordetella parapertussis and Bordetella bronchiseptica”. Nature Genetics. 35 (1): 32–40. doi:10.1038/ng1227. PMID 12910271.
- Goldman, W.E. and J.B. Baseman (March 1980). “Abstr. Ann. Meet. Am. Soc. Microbiol”. Abstr. Ann. Meet. Am. Soc. Microbiol.[page needed]
- Goldman, William E.; Klapper, David G.; Baseman, Joel B. (1982-05-01). “Detection, isolation, and analysis of a released Bordetella pertussis product toxic to cultured tracheal cells”. Infection and Immunity. 36 (2): 782–94. doi:10.1128/iai.36.2.782-794.1982. PMC 351298. PMID 6177637.
- Cookson, BT; Tyler, AN; Goldman, WE (1989). “Primary structure of the peptidoglycan-derived tracheal cytotoxin of Bordetella pertussis”. Biochemistry. 28 (4): 1744–9. doi:10.1021/bi00430a048. PMID 2541765.
- Wilson, R; Read, R; Thomas, M; Rutman, A; Harrison, K; Lund, V; Cookson, B; Goldman, W; et al. (1991-01-01). “Effects of Bordetella pertussis infection on human respiratory epithelium in vivo and in vitro”. Infection and Immunity. 59 (1): 337–45. doi:10.1128/iai.59.1.337-345.1991. PMC 257746. PMID 1987048.
- Luker, K. E.; Collier, J. L.; Kolodziej, E. W.; Marshall, G. R.; Goldman, W. E. (1993). “Bordetella pertussis Tracheal Cytotoxin and Other Muramyl Peptides: Distinct Structure-Activity Relationships for Respiratory Epithelial Cytopathology”. Proceedings of the National Academy of Sciences. 90 (6): 2365–2369. Bibcode:1993PNAS…90.2365L. doi:10.1073/pnas.90.6.2365. PMC 46087. PMID 8460147.
- Jacobs, C; Joris, B; Jamin, M; Klarsov, K; Van Beeumen, J; Mengin-Lecreulx, D; Van Heijenoort, J; Park, JT; et al. (1995). “AmpD, essential for both beta-lactamase regulation and cell wall recycling, is a novel cytosolic N-acetylmuramyl-L-alanine amidase”. Molecular Microbiology. 15 (3): 553–9. doi:10.1111/j.1365-2958.1995.tb02268.x. PMID 7783625. S2CID 40169153.
- Wilson, R; Read, R; Thomas, M; Rutman, A; Harrison, K; Lund, V; Cookson, B; Goldman, W; et al. (1991). “Effects of Bordetella pertussis infection on human respiratory epithelium in vivo and in vitro”. Infection and Immunity. 59 (1): 337–45. doi:10.1128/iai.59.1.337-345.1991. PMC 257746. PMID 1987048.
- Swaminathan, C. P.; Brown, P. H.; Roychowdhury, A.; Wang, Q.; Guan, R.; Silverman, N.; Goldman, W. E.; Boons, G.-J.; Mariuzza, R. A. (2006). “Dual strategies for peptidoglycan discrimination by peptidoglycan recognition proteins (PGRPs)”. Proceedings of the National Academy of Sciences. 103 (3): 684–9. Bibcode:2006PNAS..103..684S. doi:10.1073/pnas.0507656103. PMC 1334652. PMID 16407132.
- Melvin, Jeffrey A.; Scheller, Erich V.; Miller, Jeff F.; Cotter, Peggy A. (2014-04-01). “Bordetella pertussis pathogenesis: current and future challenges”. Nature Reviews Microbiology. 12 (4): 274–288. doi:10.1038/nrmicro3235. ISSN 1740-1526. PMC 4205565. PMID 24608338.
- Coote, JG (2001). Environmental sensing mechanisms in Bordetella. Advances in Microbial Physiology. Vol. 44. pp. 141–81. doi:10.1016/S0065-2911(01)44013-6. ISBN 9780120277445. PMID 11407112.
- Swaminathan, C. P.; Brown, PH; Roychowdhury, A; Wang, Q; Guan, R; Silverman, N; Goldman, WE; Boons, GJ; Mariuzza, RA (2006). “Dual strategies for peptidoglycan discrimination by peptidoglycan recognition proteins (PGRPs)”. Proceedings of the National Academy of Sciences. 103 (3): 684–9. Bibcode:2006PNAS..103..684S. doi:10.1073/pnas.0507656103. PMC 1334652. PMID 16407132.
- Humann, Jessica; Lenz, Laurel L. (2009). “Bacterial Peptidoglycan-Degrading Enzymes and Their Impact on Host Muropeptide Detection”. Journal of Innate Immunity. 1 (2): 88–97. doi:10.1159/000181181. PMC 2659621. PMID 19319201.
- Magalhaes, Joao Gamelas; Philpott, Dana J; Nahori, Marie-Anne; Jéhanno, Muguette; Fritz, Joerg; Bourhis, Lionel Le; Viala, Jérôme; Hugot, Jean-Pierre; et al. (2005). “Murine Nod1 but not its human orthologue mediates innate immune detection of tracheal cytotoxin”. EMBO Reports. 6 (12): 1201–7. doi:10.1038/sj.embor.7400552. PMC 1369207. PMID 16211083.
- Skerry, C; Goldman, WE; Carbonetti, NH (February 2019). “Peptidoglycan Recognition Protein 4 Suppresses Early Inflammatory Responses to Bordetella pertussis and Contributes to Sphingosine-1-Phosphate Receptor Agonist-Mediated Disease Attenuation”. Infection and Immunity. 87 (2). doi:10.1128/IAI.00601-18. PMC 6346131. PMID 30510103.
- Chang, CI; Chelliah, Y; Borek, D; Mengin-Lecreulx, D; Deisenhofer, J (24 March 2006). “Structure of tracheal cytotoxin in complex with a heterodimeric pattern-recognition receptor”. Science. 311 (5768): 1761–4. Bibcode:2006Sci…311.1761C. doi:10.1126/science.1123056. PMID 16556841. S2CID 25416989.
External links
- Ligand entry in the Protein Data Bank: MLD
- Entry in MetaCyc: CPD0-1080
Leave a Reply